

1. 서 론
2. 조사지 개황
3. 연구방법
3.1 식생 구조 조사
3.2 식생도 작도
3.3 자료 처리
4. 결과 및 논의
4.1 식생 종류
4.2 식생 구조
4.3 식생 분포
5. 결론 및 제언
1. 서 론
우리나라에서는 신석기시대인 기원전 4,000년부터 논 농사가 성행하여 왔고 이와 관련된 수리관개시설 유물이 다수 잔존하여 있다 (Heu and Moon 2010). 특히 삼한시대에 축조된 밀양 수산제, 김제 벽골제, 제천 의림지 등이 현재까지 남아있다 (Hwang et al. 2014). 본 연구의 대상지인 공검지는 통일신라시대 후기인 8세기에 축조되었고, 그 후 기후변화와 저수지 내 퇴적에 의하여 수 차례 제방을 증축하여 왔다 (Hwang et al. 2014). 고려시대에 공검지는 제방의 길이 430 m, 저수지 둘레 8.7 km, 최대수심 5.6 m로 추정되어 수심은 얕지만 거대한 면적의 고대 저수지이었다고 추정되고 있다 (Yoon et al. 2013, Hwang et al. 2014). 일제 강점기 이후에 대부분의 저수지가 농경지로 개간되고 극히 일부의 수면만이 잔존하여 현재는 저수지 둘레가 단지 1.3 km에 지나지 않아서 과거 둘레의 1/7 수준으로 축소되었다. 현재 지형도로 보면 저수지 제방 상류의 유역이 고도가 낮아서 넓은 면적에 걸쳐 저수지를 만들기에 용이한 지역임을 알 수 있다.
경상북도 상주시 공검면에 위치한 공검지는 경상북도 기념물 제121호로 지정되어 있다. 또한 공검지는 역사, 문화적 가치뿐만 아니라 생태적 가치가 높아서 환경부는 2011년 6월 이곳을 국내 최초로 논 습지보호지역으로 지정하였다 (DREO 2014). 공검지 습지보호지역이 특이한 것은 이 보호지역 안에 저수지 습지뿐만 아니라 산림, 하천, 농경지가 포함되어 있다는 것이다. 따라서 이 보호지역은 습지의 현명한 이용에 대한 좋은 본보기가 될 수 있는 조건을 갖춘 것으로 생각된다 (Finlayson et al. 2011).
공검지에 대한 생태조사 결과를 살펴보면, 대구지방환경청에서 습지보호지역의 보전을 위하여 발행한 종합 조사보고서가 있다 (DREO 2014, 2017, 2018). 또한 2012년에 실시한 식물상 조사에 따르면 이 보호지역에서 총 240분류군이 발견되었고, 이중 수생식물이 31분류군이었는데 이는 우리나라의 수생식물 총종류의 18%에 해당하였다 (Song and Park 2013). 곤충에 대한 연구에서는 2014년에 총 227종의 곤충을 보고하여 이 습지가 비교적 안정적인 환경에 있다고 하였다 (Kim et al. 2017). 한편 고생태학적 연구로서 퇴적토에서 규조류의 변화를 조사하여 저수지 과거 환경을 추정한 연구에서 저수지의 역사적 변천과 기후변화를 추론하였다 (Hwang et al. 2014, Lee et al. 2018). 또한 퇴적층에 대한 지질학적 연구에 의하여 저수지의 기후변화에 대하여 조사하였다 (Yoon et al. 2013). 그러나 이곳에서 식생 구조와 분포에 대하여 조사한 보고서 이외의 학술 보고는 아직 없다.
따라서 본 조사에서는 공검지 습지보호지역의 지속가능한 보전을 위한 전략을 수립하고 생태계 변화를 추적하기 위한 기초자료를 수립하기 위하여 1) 습지와 인접 육상지역에서 나타나는 현존 식생을 분류하고, 2) 분류된 식생의 구조적 특징을 규명하고, 3) 이들 식생의 분포지를 조사하여, 4) 최종적으로 생태계 보전을 위한 제언을 하고자 한다.
2. 조사지 개황
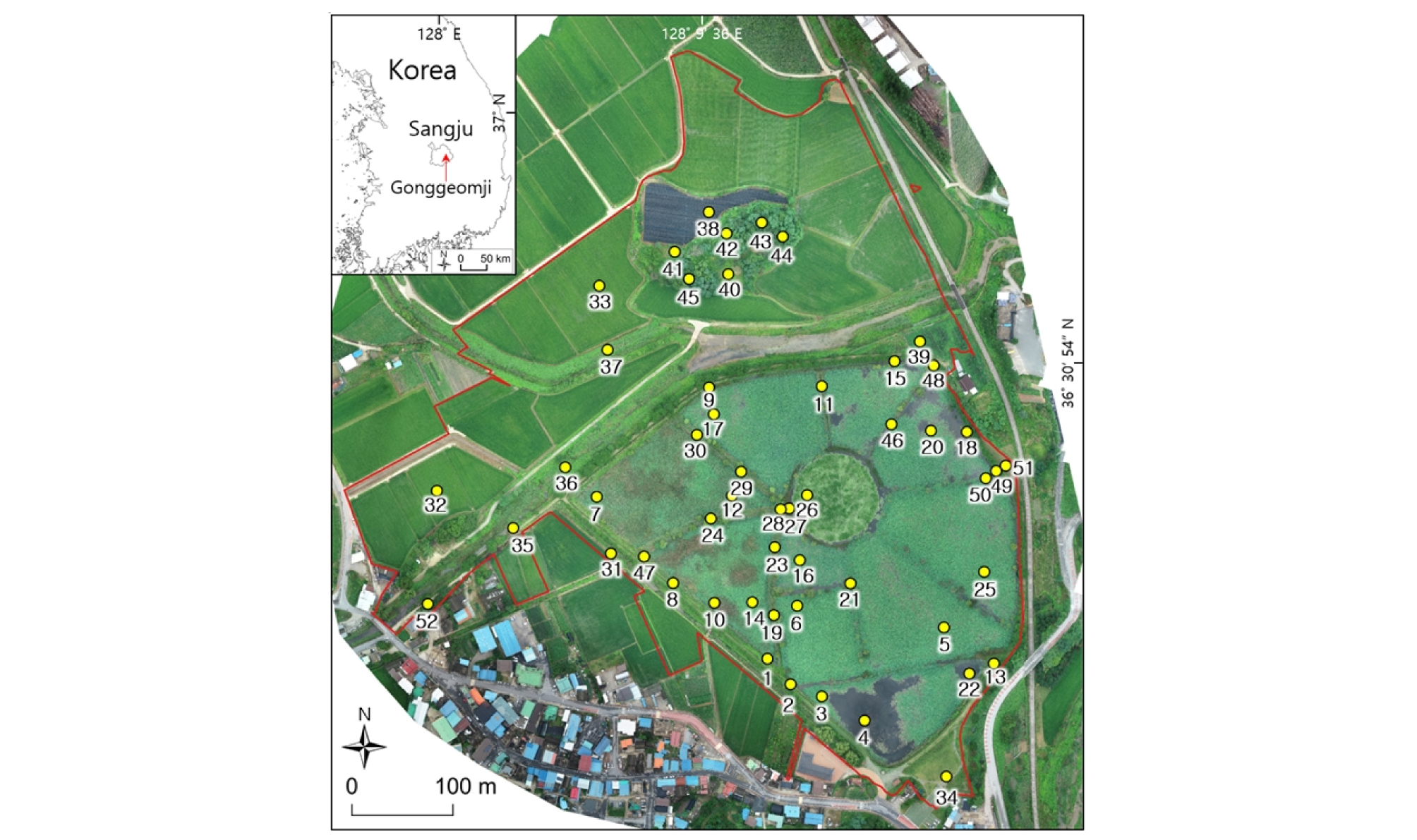
본 연구의 조사지는 경상북도 상주시 공검면 양정리에 위치한 공검지 습지보호지역이다 (Fig. 1). 보호지역의 총면적은 26.2 ha이고 이중에서 공검지 습지에 해당하는 면적은 14.7 ha (56%)이었다. 공검지는 수심이 얕은 저수지 습지 유형이다. 공검지 습지보호지역은 저수지뿐만 아니라 주변의 하천, 산림, 농경지 등의 다른 토지이용 지역도 포함하고 있다. 주요 지역의 개황은 다음과 같다.
1) 공검지 저수지: 저수지 습지가 공원용으로 정형적인 구획이 나누어져 있다. 특히 중앙부에는 원형의 습지가 조성되어 있고 이곳을 중심으로 방사상으로 구역이 나누어져 있다. 각 구획의 경계를 따라서 성토한 길이 축조되어 있다. 이 습지로의 유입수는 습지의 북서쪽에서 농수로에 연결된 개수로의 수문을 통하여 유입되고 있다. 또한 유출수는 습지의 남쪽 끝에서 수문을 통하여 암거로 유출되고 있다. 습지의 수심은 유입수 부근에서 얕고 유출수 부근과 저수지의 북동쪽이 깊은 편이었다.
2) 동천 및 농수로: 공검지 습지의 북쪽 주변을 따라서 동천이 서쪽에서 동쪽으로 흐르고 있다. 습지와 동천은 제방으로 나누어져 있으며 둑마루를 비롯한 습지의 가장자리를 따라서 차량이 운행할 수 있는 도로가 축조되어 있다. 두 하천이 합류하는 하류에는 최근 하천정비 공사에 의하여 제방사면과 저수로가 새롭게 형성되었다. 한편 공검지의 남서쪽 경계를 따라서 제방과 연결하여 농수로가 흐르고 있다.
3) 산림 구릉지: 습지보호지역 중앙부의 북쪽에 작은 구릉의 산지가 잔존하고 있다. 이곳에는 산림, 묘지 초지 및 인삼 밭이 위치하고 있다.
4) 농경지 및 과수원: 공검지의 북쪽 지역에는 넓게 논이 둘러 싸고 있다. 또한 부분적으로 콩 밭, 밤나무 및 사과나무 과수원이 조성되어 있다. 그리고 습지 관리사무소와 역사박물관이 위치한 남쪽 출입구에는 잔디밭이 조성되어 있다.
조사지 전역에서 식생의 상관, 서식처 유형과 습지의 수심을 고려하여 총 52개의 조사지점을 선정하였다 (Fig. 1). 특히 저수지 습지와 산림 지역을 중점조사지로 선정하여 이곳에 대하여 세밀한 식생조사를 실시하였다. 습지에 39개, 산지에 6개, 조림지에 2개, 경작지에 4개 및 잔디밭에 1개의 조사 방형구를 설치하여 식생을 조사하였다.
공검지에서 식생 조사는 총 3회 실시하였다. 1차 조사는 2019년 5월, 2차 조사는 7월 및 3차 조사는 9월에 실시하였다. 특히 2차 조사기에 식생구조와 식생도 조사를 집중적으로 실시하였다.
3. 연구방법
3.1 식생 구조 조사
식물군집 조사는 전통적인 식물사회학적 방법을 따랐다 (Braun-Blanquet 1965, Kim and Lee 2006). 현장에서 식생조사구의 위치 선정은 해당 서식처의 상관, 종조성 및 환경을 종합적으로 고려하여 그 서식처를 대표할 수 있는 전형적인 종조성과 균질한 환경을 갖춘 곳을 택하였다. 식생 조사구의 면적은 원칙적으로 식생 최고 높이의 자승한 값에 준하는 면적을 정하였다. 즉, 부수, 침수, 부엽 식생, 논과 밭, 습초지 및 건초지에서는 1 m x 1 m, 정수식생에서는 2 m x 2 m, 버드나무림과 과수원에서는 5 m x 5 m, 교목 산림 식생에서는 10 m x 10 m의 방형구를 설치하였다. 출현 식물종의 정량적 평가는 Westhoff and van der Maarel (1973)가 Braun-Blanquet 척도를 변환한 피도-우점도 척도를 이용하였다. 한편 식생을 조사할 때 습지에서 가장 중요하다고 생각되는 환경요인인 수심을 자를 이용하여 측정하였다.
3.2 식생도 작도
공검지 습지보호지역에서 식생의 분포를 파악하기 위하여 현존식생도를 작도하였다. 식생도 작도를 위하여 DJI 사의 Phantom 4 pro를 이용하여 공중 영상을 촬영하였다. 영상을 촬영할 때는 Map Pilot for DJI – Business 어플리케이션을 이용하여 100 m 상공에서 지상 사진 영상이 75 % 겹치도록 촬영 간격을 조정하였다. 촬영된 영상은 Pix4D 사의 PIX4Dmapper를 이용하여 정사 보정한 영상으로 지도화하였다. 식생도를 작도하기 위하여 정사 보정된 영상을 인쇄한 사진에 분류된 식물군집의 경계를 현장에서 눈으로 관측하여 그렸다. 현장에서 작도한 식생의 경계를 QGIS (ver 3.4)에 입력하여 현존식생도를 작도하였다. 각 식물군집의 분포면적은 QGIS (ver 3.4)에서 계산하였다.
3.3 자료 처리
야외에서 조사한 식생 자료를 종합하여 식물사회학의 표 작업으로 식물 군집을 분류하였다 (Kim and Lee 2006). 분류된 식물군집의 이름은 해당 군집의 우점종명을 이용하여 명명하였다. 또한 식생 자료를 활용하여 다양한 자료 및 통계 처리를 수행하기 위하여, 식물사회학적 조사에 의하여 구하여진 피도-우점도 척도를 Van der Maarel (1979)의 방법에 따라서 피도로 전환하였다.
우선 각 방형구에서 출현종별 전환된 피도를 이용하여 상대피도를 산출하여 우점종과 기타 피도가 높은 종을 선별하였다. 또한 각 식물군집의 종다양성을 비교하기 위하여 Shannon index를 계산하였다 (Shannon 1948). 한편 식생 구조와 식생과 환경과의 관계는 다변량 분석인 탈경향대응분석 (detrended correspondence analysis, DCA)으로 분석하였다. 종별 피도 자료를 로그 변환하여 R ver. 3.5.1 (R Core Team 2019) 환경에서 vegan 패키지의 “decorana” 함수로 DCA를 수행하였다 (Oksanen 2019). 환경요인인 수심과 식생과의 관계를 파악하기 위하여 vegan의 “envfit” 함수를 이용하여 분석하였다 (Oksanen 2019). 마지막으로 식물종 특성과 식생구조의 관계를 구명하기 위하여 식물종 특성을 생활형에 따라서 침수식물, 부수식물, 부엽식물, 정수식물, 습생식물, 육상식물로 구분하였고, 또한 외래종 여부에 따라서 고유종과 외래종으로 구분하였다. 식물종 특성의 분류는 Jeong et al. (2012)를 참고하였다. DCA 분석을 위한 식물종 특성 자료는 각 방형구에서 해당 생활형 또는 외래종의 종 특성의 빈도를 구하여 만들었다. 이러한 종 특성 자료를 환경요인 분석과 같은 방법으로 vegan의 “envfit” 함수를 이용하여 분석하였다 (Oksanen 2019).
4. 결과 및 논의
4.1 식생 종류
식물사회학적 방법으로 공검지 습지보호지역에서 총 26개 식물군집으로 분류하였다 (Table 1). 본 조사지역은 삼림, 농경지, 하천 및 습지 등을 포괄하고 있기 때문에 매우 이질적이고 다양한 식물군집이 출현하였다. 생활형 및 상관에 따라 구분한 군집은 다음과 같다.
1) 부수침수 식생: 참통발 군집, 검정말 군집, 마름-붕어마름 군집이 속하는데 참통발은 부수성, 검정말과 붕어마름은 침수식물이지만 뿌리가 끊어져도 생존이 가능하며, 마름은 부엽식물이지만 뿌리 없이 부수할 수 있다 (Shalabh et al. 2012).
2) 부엽 식생: 수련 군집이 여기에 속하였다. 공검지의 수련 군집은 조경용 종이 식재된 것으로 생각된다. 부수침수 및 부엽 식물군집은 부수성 또는 부엽성 특성을 가지고 있기 때문에 수질이 오염된 부영양 습지에서 탁도가 높은 물 위를 차지하여 다른 식물에 대하여 경쟁력을 가질 수 있다 (Hough et al. 1989).
3) 정수 식생: 이 식생은 주로 공검지 습지에 분포하는 연꽃, 큰고랭이, 큰잎부들, 애기부들, 갈대가 우점하는 군집이었다. 일반적으로 이들 군집은 안정된 습지에서 한 종이 과우점하는 단일종 군집을 형성하는데 반하여 (Daoust and Childers 1999), 공검지에서는 다양한 종이 혼재하는 정수식물 군집을 형성하는 것이 특이하였다. 정수 식생 중에서 줄과 갈대가 우점하는 군집은 주로 동천과 농수로의 흐르는 물에 분포하였다. 공검지에서 정수식물 군집이 발달하여 있는 것은 습지의 육역화와 부영양화가 진행되면서 정수식물 군집이 우세하게 된 것으로 생각된다 (Vaithiyanathan and Richardson 1999).
4) 습생 식생: 습초지 군집으로서 여뀌와 명아자여뀌가 우점하는 군집이 속하였고 지형이 약간 높아서 수심은 없으나 수분으로 포화된 땅에서 발견되었다.
5) 나무습지 (Swamp) 식생: 버드나무-갈대 군집이 속하며, 버드나무 수관 아래 갈대가 수면 또는 지면을 덮고 있었다.
6)건생초지 식생: 습지보호지역의 중앙 상부에 위치한 산지에서 발견된 군집으로서 묘지 식생인 띠-조개나물 군집이 여기에 속하였다.
7) 하록활엽수림 식생: 산지에서 발견된 군집으로서 아까시나무가 혼생하는 상수리나무와 갈참나무 군집이 교목림을 형성하고 있었다.
8) 관리 교란 식생: 농경지로서 논, 콩, 인삼 군집과 과수원으로서 밤, 사과나무 군집이 분류되었다. 또한 조경용으로 조성된 잔디 군집이 여기에 속하였다.
Table 1. Dominant and abundant species with their relative cover and distribution area of the plant communities of the Gonggeomji Wetland Protection Area, South Korea
4.2 식생 구조
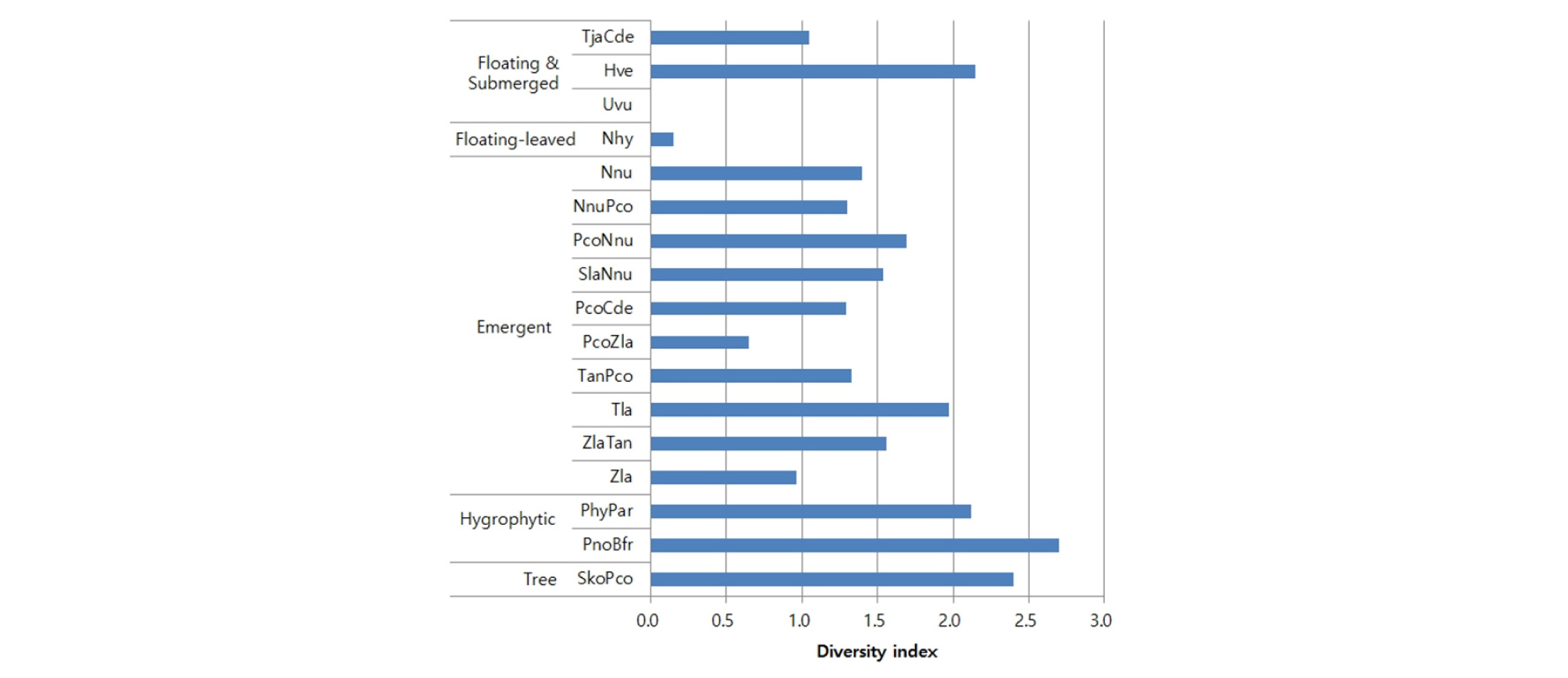
공검지 습지보호지역에서 나타나는 식물군집의 Shannon 종다양성지수는 군집에 따라서 매우 큰 차를 나타내었다 (Fig. 2). 저수지 습지에서 종다양성이 낮은 군집은 참통발 군집이 비교적 수심이 깊은 개방수면에서 단일종이 출현하였고, 수련 군집이 과밀한 부엽에 의한 수면을 피압하였으며, 대부분의 정수식물 군집이 한두 종이 우점하는 특성을 나타내었다. 반면에 종다양성이 높은 군집은 습생식물 군집이 습초지에서 적당히 교란이 가해지는 곳으로서 중간교란가설에 의하여 높은 종다양성이 설명이 되는 것으로 생각된다 (Wilkinson 1999, Roxburgh et al. 2004). 또한 버드나무 군집은 비교적 지형이 높고 목본과 초본 식물이 혼생하여 종다양성이 높은 군집으로 유지되고 있었다. 습지 식생 이외 군집의 종다양성은 띠-조개나물 군집 2.46, 상수리나무-아까시나무 군집 1.89, 갈참나무-아까시나무 군집 2.79이었다. 한편 관리지역 군집의 종다양성은 밤나무 군집 1.74, 사과나무 군집 1.32, 벼 군집 0.39, 인삼 군집 0, 콩 군집 0.55, 잔디 군집 0.93으로서 과수원이 농경지 군집보다 높았다.
공검지 습지 식생간의 관계와 식생과 환경 사이의 관계를 분석하기 위하여 적용한 탈경향대응분석 (DCA)에서 1축과 2축의 설명력이 각각 40% 및 25%으로 총 65%에 달하여 이 두 축으로 전체 식생 분포 경향을 비교적 잘 파악할 수 있었다 (Fig. 3). 군집의 배열에서 1축의 왼쪽에 수련, 연꽃 등의 군집이 오른쪽에 습초지 군집이 위치하고 있었고 왼쪽 아래로 참통발, 마름-붕어마름 군집 등이 배열되었다 (Fig. 3 (a)). 종의 배열에서는 왼쪽 아래에 마름, 참통발 등의 부수침수 식물종이 오른쪽 위쪽에 교란지에 발생하는 명아자여뀌, 여뀌, 미국가막사리 등이 위치하였다 (Fig. 3 (b)). 환경요인 중에서 수심이 왼쪽 아래로 갈수록 깊어져서 식물군집의 배열 경향을 잘 설명하고 있었다 (Fig. 3 (c)). 마지막으로 식물종 특성을 살펴보면 1축의 오른쪽에 습지에서 교란지 특성을 나타내는 외래종과 육상식물이 위치하였으며 2축의 위에 정수식물이 아래에 침수식물이 배치되어 수심에 따른 대형수생식물의 분포 경향을 잘 설명하고 있었다 (Fig. 3 (d)). 이러한 배열 특성을 종합하면, 공검지 습지에서 식물군집이 교란 정도와 수심 환경경사에 따라서 분포한 것으로 생각된다. 일반적으로 습지 식생에서 수심이 절대적인 영향인자이지만 (Miller and Zedler 2003), 공검지에서는 인간이나 범람에 의한 교란이 중요한 환경요인으로 작용하였다 (Bornette and Amoros 1996, Tousignant et al. 2010).
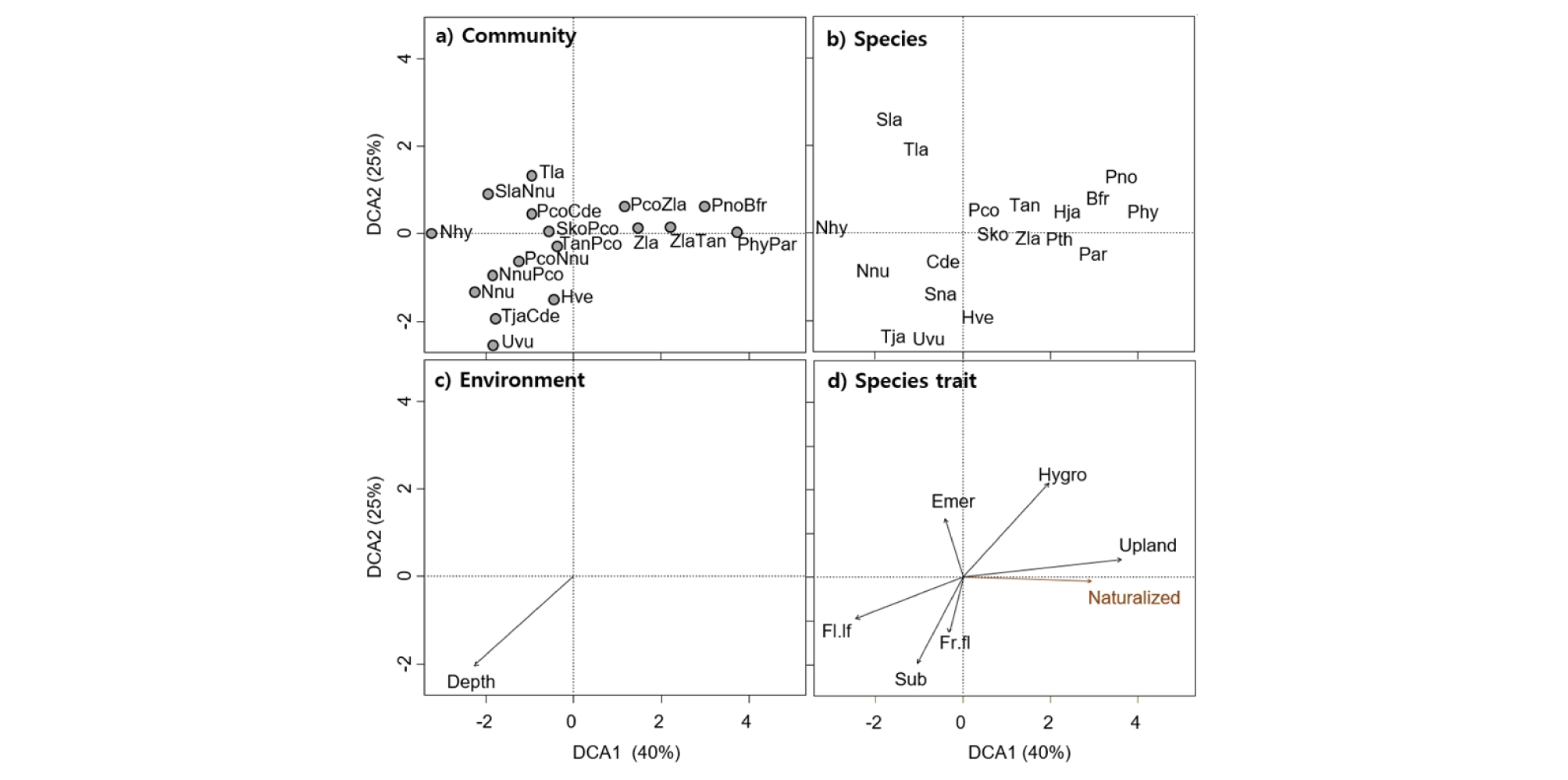
Fig. 3.
Biplots of detrended correspondence analysis (DCA). (a) Plant community (abbreviation refers to Table 1), (b) Plant species (abbreviation refers to Table 1 and Hja = Humulus japonicus, Pth = Persicaria thunbergii, Sna = Salvinia natans), (c) Environmenal factor (the arrow indicates the direction and magnitude of water depth), (d) species traits (Emer = emergent macrophytes, Fl.lf = floating-leaved macrophytes, Fr.fl = free-floating macrophytes, Hygro = hygrophytes, Naturalized = naturalized species, Sub = submerged macrphytes, Upland = upland plants).
4.3 식생 분포
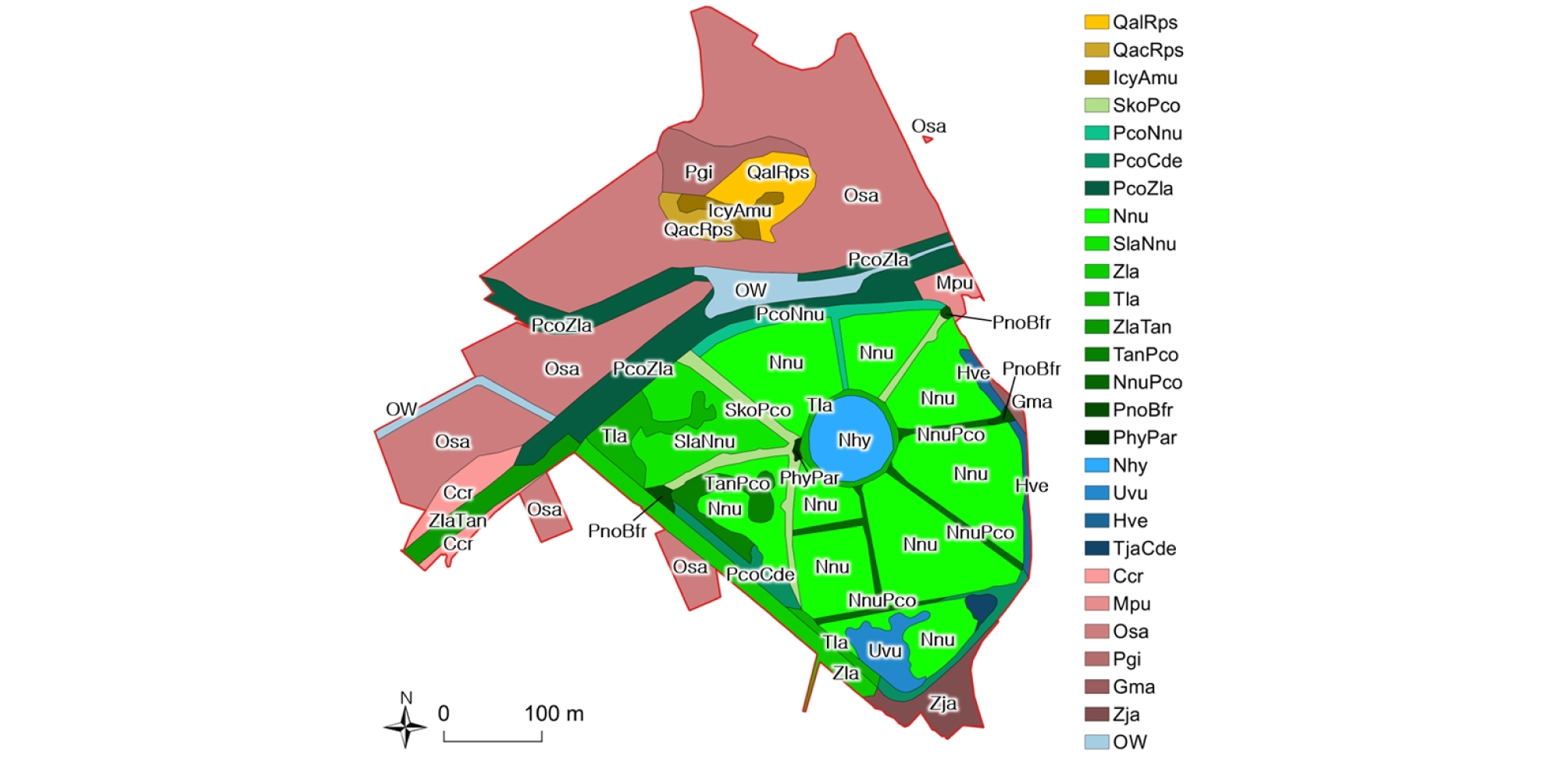
공검지 습지보호지역에서 서식처 유형별로 식물 군집의 분포를 살펴보면 다음과 같다 (Fig. 4). 저수지 습지에는 주로 정수식생이 분포하고 일부 수역에 부수침수 식생, 나무습지 식생 및 습생 식생이 분포하였다. 구획으로 나누어진 습지의 중앙 원형 구획에는 수련 군집이 분포하였다. 나머지 구획의 대부분은 연꽃 군집과 연꽃을 포함한 정수식물 군집이 차지하고 있었다. 수심이 깊은 곳에 참통발과 침수식물 군집이 습지의 남쪽과 동쪽 일부 수역을 차지하고 있었다. 습지의 북서쪽에 정수식물 군집이 위치하며 지형이 높은 제방 사면과 구획을 나누는 길 위에는 버드나무 군집이 위치하였다. 하천 식생으로서 동천과 농수로에는 줄 및 기타 정수식물이 혼생하는 군집이 넓은 수면과 홍수터를 덮고 있었다. 단지 동천과 지류가 합류되는 곳에만 개방수면이 있었다. 한편 산지에는 하록활엽수림 식생인 상수리나무-아까시나무, 갈참나무-아까시나무 군집이 위치하며 그 중간에 묘지에 건생이차초원 군집이 분포하였다. 농업 및 관리지 식생으로서 논, 밭, 과수원 그리고 길가 식생이 공검지 습지의 북쪽과 남쪽에 분포하였다.
전체 습지보호지역 내 군집 별 분포 면적은 습지의 수생식물 식생 면적 (전체 지역의 48%)이 가장 넓고 농경지인 논의 면적 (32%)도 넓었다. 하천을 포함한 습지에서 한정하여 분포면적을 살펴보면, 연꽃 군집 (습지의 46%)이 가장 분포면적이 넓었다. 다른 식물과 혼생하는 연꽃의 총 분포면적은 8.4 ha (60%)를 차지하여 절반 이상의 면적을 차지하고 있었다. 하천에는 갈대-줄 군집이 넓은 면적을 차지하였다.
공검지 습지의 2014년 이후 식생분포의 경년변화를 살펴보면, 식생이 점차 육역화, 단순화되고 있었다 (DREO 2014, 2017, 2018). 즉 개방수면과 부수식물인 생이가래 군집의 분포지가 감소하였고, 정수식물의 분포 면적이 증가하고 있었다. 또한 2017년 이후에 물 관리의 변화로 수위가 상승하여 갈대와 버드나무 군집이 감소하였다. 또한 연꽃이 급격하게 분포면적을 확장하고 있었는데, 이는 연꽃의 지하경에 의하여 빠르게 번식하고 수면을 메워서 다른 식물을 피압하기 때문이다. 한편 애기부들, 큰잎부들 등의 정수식물의 분포 면적이 증가하고 있다. 습지의 북서쪽 수변부를 중심으로 점차 정수식물이 확장되고 있는데, 이는 이 부근에서 유입수가 유입되고 있고 유입수가 유입되면서 이곳에 토사가 퇴적되어 수심이 점차 얕아지면서 정수식물이 확장되고 있는 것으로 판단된다. 습지의 북쪽에 위치한 동천에는 줄이 급속하게 정착 및 확장되어 하천을 덮고 있었다. 이러한 이유는 동천 상류에 위치한 오태저수지에 의하여 하류 하천의 유량이 조절되기 되어 홍수에 의한 교란이 감소하였거나 영양물질의 유입량이 증가하여 식물의 생장이 촉진되기 때문인 것으로 생각된다 (Yi et al. 2019, Woo et al. 2019).
5. 결론 및 제언
공검지 습지보호지역에서 총 26개 식물군집이 분류되었는데, 저수지 습지, 하천, 산지, 농경지 등의 이질적인 서식처에서 다양한 식물군집이 분포하였다. 주요 서식처의 식생 현황과 이에 따른 관리방안을 살펴보면 다음과 같다. 먼저, 식충식물인 참통발 군집은 수심이 깊은 유출수 부분의 개방수면에 주로 분포하였다. 희귀종인 참통발을 보전하기 위하여 연꽃 등의 부엽식물의 침입을 제한하여 개방수면을 확보하는 것이 필요하다고 생각된다. 현재 수련 군집은 습지 중앙부에 식재되어 수면을 완전히 피복하고 있다. 습지 중앙부는 격리된 핵심지역으로서 자연식생으로 복원이 필요하다고 생각된다. 저수지에서 분포 면적이 가장 넓은 연꽃 군집은 습지의 대부분 지역이 수심 0.5 - 2 m로 얕아서 연꽃이 과도하게 확장하고 있었다. 습지에 퇴적을 억제하여 깊은 수심을 확보하고 연꽃 지하경의 확장을 억제할 수 있는 장벽을 저토에 설치할 필요가 있다. 큰잎부들, 애기부들-갈대 등의 정수식물 군집은 습지의 가장자리를 따라서 발달하여 있다. 부들류로 식생이 단순화되고 있으므로 줄, 큰고랭이, 세모고랭이, 창포 등의 다양한 정수식물을 유도하여야 한다고 생각된다. 여뀌-갈풀, 명아자여뀌-미국가막사리 군집은 비교적 지형이 높은 교란된 습생초지에 발달하여 있었다. 이곳으로 외래종의 유입을 차단하고 향유, 박하 등의 고유 식생이 정착할 수 있도록 관리하여야 한다. 마지막으로 하천 식생으로서 동천에는 줄, 갈대-줄 군집 등의 식생이 과번무하여 단순하고 생태계교란종인 환삼덩굴이 침입하여 있다. 생태하천 복원 기법을 도입하여 공검지 습지와의 연계성을 확보하는 것이 필요하다고 생각된다.
식생 조사 결과를 종합하여 공검지 습지보호지역의 지속가능한 보전과 복원을 위하여 다음 사항을 제언한다.
1) 수문 체계의 회복과 관리: 유입수, 유출수 및 습지 내부의 물 순환 체계를 파악하여 습지 전체로 물 순환이 원활하도록 수문 체계를 회복하여야 한다. 현재는 유입수가 습지 전체로 흐르지 못하고 주로 저수지의 서쪽을 따라서 흐르고 있는 것으로 판단된다. 한편 현재 수심과 물 흐름을 조절하기 위하여 유입수의 관리를 관리자가 임의로 하고 있다. 공검지의 생태계 기능과 구조를 유지하기 위하여 필요한 수문 관리 요령을 지침서로 작성하여 이에 따라서 운영하도록 하여야 한다.
2) 육역화 방지: 현재 부엽식물, 정수식물, 습생식물이 저수지 전체로 확대되어 육역화가 빠르게 진행되고 있다. 우선 습지로 물이 유입되기 전에 침사지를 설치하여 토사가 유입되는 것을 막아야 한다. 또한 유입수에 의하여 유입되는 토사가 많은 시기에는 물 관리를 통하여 유입수량을 제한하여 토사 유입량을 줄여야 할 것으로 생각된다.
3) 개방수면의 확대: 현재 저수지 수면이 수생식물에 의하여 거의 피복된 상태로서 다른 생물의 다양성 확보를 위하여 전체 면적의 약 절반 정도를 개방수면으로 유도할 수 있도록 충분한 수심을 확보하고 지형을 조성하야 한다.
4) 지형과 식생의 자연화: 현재 조경, 공원 용도 구획화되어 변형된 인위적인 지형을 자연 습지로 복원하여야 한다. 또한 현재 조경용으로 식재된 수련, 연꽃 등의 식생을 우리나라 고유의 식생으로 유도하여야 한다. 특히 연꽃은 왕성한 지하경 성장으로 과도하게 저수지를 덮고 있다. 따라서 창포, 올방개, 털부처꽃 등의 전통 저수지 환경에 알맞은 우리 고유의 다양한 식생으로 유도할 필요가 있다.
5) 공간 구획화와 연결성 회복: 습지보호지역을 핵심 지역, 완충 지역, 친수 지역으로 구분하여 관리대책을 수립하여야 한다. 특히 보호지역에 인접한 인가, 과수원, 콩밭 등은 자연지역으로 회복하여 외부 교란의 유입을 차단하여야 할 것으로 생각한다. 한편 습지보호지역에서 저수지 습지, 하천, 논, 산림으로 연결되는 생태적 연결성을 확보하여야 한다. 현재 보호지역의 관리가 주로 저수지에서만 이루어지고 있다. 관리 지역을 다른 생태계까지 확장하여 하천복원, 친환경 농업, 산림 자연화를 시도하여 완전히 연결되는 보호지역으로 구성하여야 한다.
