

1. 서 론
2. 하천생물지형학의 이해
2.1 생물과 지형 상호작용의 시간적 변화
2.2 생물과 지형 상호작용의 공간적 변이
2.3 수문지형의 이질성과 생물다양성의 관계
2.4 교란과 생물지형 복합체의 관계
3. 우리나라 하천에서 수문지형과 생물 교란
3.1 식생 번무화
3.2 외래식물 침입
4. 결론 및 전망
1. 서 론
생물지형학 (biogeomorphology)은 지형계와 생태계 사이의 상호작용을 연구하는 학문이라고 정의할 수 있다. 진화학자 다윈은 그의 저서 “The Formation of Vege Mould, Through the Action of Worms, with Observations on Their Habits”에서 생물을 최초로 지형 매개자로서 제안하였다 생물지형 복합체는 지형, 선구, 생물지형 및 생태적 단계의 순서로 전환이 된다. 공간적으로 물 흐름과 유사 분포가 식생과 상호작용하여 하도 지형의 변화가 일어난다. 이렇게 형성된 하천의 공간적 이질성은 하안의 생물종다양성을 증가시킨다. 그러나 댐 하류 하천에서는 서식처 유형과 조건이 심각하게 훼손되어 생물다양성이 저하된다. 우리나라의 하천에서는 최근 국지적 교란과 전 지구적 기후변화로 하안 식생이 번무하고 특히 외래종이 빠르게 정착하고 있다. 따라서 급격한 기후변화와 인류에 의한 압박의 시대에서 하천 생물과 수문지형 조건 사이의 상호 관계를 이해하는 것이 더욱 중요할 것으로 생각된다. 이 총설에서 논의한 하천 생물지형 개념틀은 우리나라 하천의 생태적 관리와 복원에 기여할 것으로 기대된다. 핵심어: 교란, 수문, 지형, 다중관계, 하천복원, 식생 (Darwin 1881). 이처럼 지렁이가 토양에 미치는 영향에서 출발한 생물지형학 연구는 점차 동물, 식물, 미생물의 다양한 생물로, 그리고 암석, 지형 등의 다양한 지형계로 확장되어 왔을 뿐만 아니라, 지형계가 생물에 미치는 영향을 포함하는 상호작용 관계로 발전하여 왔다 (Shaler 1892, Cowles 1899, Sprugel 1980, Stallins 2006, Coombes 2016). 이후 생물교란 (bioturbation)의 영향, 생물기원 지형, 생물표토층 (biomantle) 형성 등을 연구한 Johnson (1990)과 식물 이외에 동물의 역할을 강조한 Butler (1999)에 의하여 생물지형학 연구가 재촉발되었다 (Stallins 2006). 최근에는 지형과 생물 사이의 상호조절 관계, 생태계와 경관의 공진화, 환경 변화와 인간 교란에 의한 관리문제를 다루는 응용생물지형학이 첨단의 생물지형학 연구 분야로 대두되고 있다 (Fei et al. 2014, Coombes 2016).
생물지형학의 한 분과인 하천생물지형학 (fluvial biogeomorphology)은 흐르는 물에 의하여 유사가 침식 또는 퇴적이 되면서 지면의 지형이 형성되고 변경되는 하천 과정에서 생물과 지형 사이의 상호작용에 대하여 연구를 한다 (Charlton 2008). 2002 - 2004년 미국의 지리학회에서 발표한 지형학 논문 중에서 40%가 하천 지형에 대한 연구였으며 (Chang and Kim 2004), 특히 최근에는 하천과 유역의 보전과 관리와 관련하여 하천생물지형학에 대한 연구가 더욱 증가하고 있다 (Viles et al. 2008, Gurnell 2012, Viles 2019).
우리나라에서는 하천의 평탄한 홍수터를 경작지, 주거지, 산업지 등으로 이용하기 위하여 하천의 수문과 지형을 변형하여 왔다. 특히 1960년대부터 하천 주변 토지이용을 극대화하고 개발된 지역을 홍수 재해로부터 보호하고 수로를 직강화하고 제방을 축조하여 하천을 개수하였다 (Lee and Yu 2001, Woo 2010, Jin et al. 2019). 또한 하천 내에서 친수공간을 확보하고 물 흐름을 집중시키기 위하여 저수로와 고수부의 복단면 구조로 하천 지형을 인위적으로 변형하는 경우도 있었다. 한편 수자원 확보와 홍수 방재를 위하여 하천의 보와 댐을 축조하여 유량을 조절하였다. 급격한 산업화 과정에서 필요한 골재를 공급하기 위하여 하천에서 모래와 자갈을 채취하여 하천 지형이 교란되어 왔다. 현재 우리나라에서는 국가하천의 96%와 지방하천 74%, 총 77%의 하천이 이미 제방축조에 의하여 하천 지형이 변형되었다 (Fig. 1, MOLIT 2019). 이와 같은 하천의 물리환경의 변화에 따라서 생물상, 생물군집 및 경관에 변화가 초래되었는데, 특히 하천의 식생번무화가 물 흐름을 방해하고 지형을 고착시키는 생물에 의한 환경 변화가 초래되었다 (Kamada et al. 2004). 이러한 하천의 생물지형 교란에 대응하여 생물다양성과 생태계서비스를 향상하기 위한 다양한 하천복원 노력이 이루어지고 있다 (Woo 2010). 따라서 몬순 기후의 우리나라에서 지속가능한 하천 관리와 복원 기술을 개발하기 위하여, 하천 생물지형에 대한 특성을 파악하고 교란에 대한 하천의 반응을 이해할 수 있는 생물지형학을 도입하는 것이 시급할 것으로 판단된다.
특히 2019년에 발족한 응용생태공학회의 하천생물지형 분과위원회에서는 하천생물지형학에 대한 소개가 선행되어야 한다는 인식을 하게 되었다. 따라서 본 총설에서는 하천생물지형학에 대한 이해를 도모하기 위하여 이 학문 분과의 정의와 연구 내용을 식생을 중심으로 살펴보고, 우리나라 하천에서 생물과 수문지형 사이의 상호작용의 대표적인 사례를 소개하고자 한다.
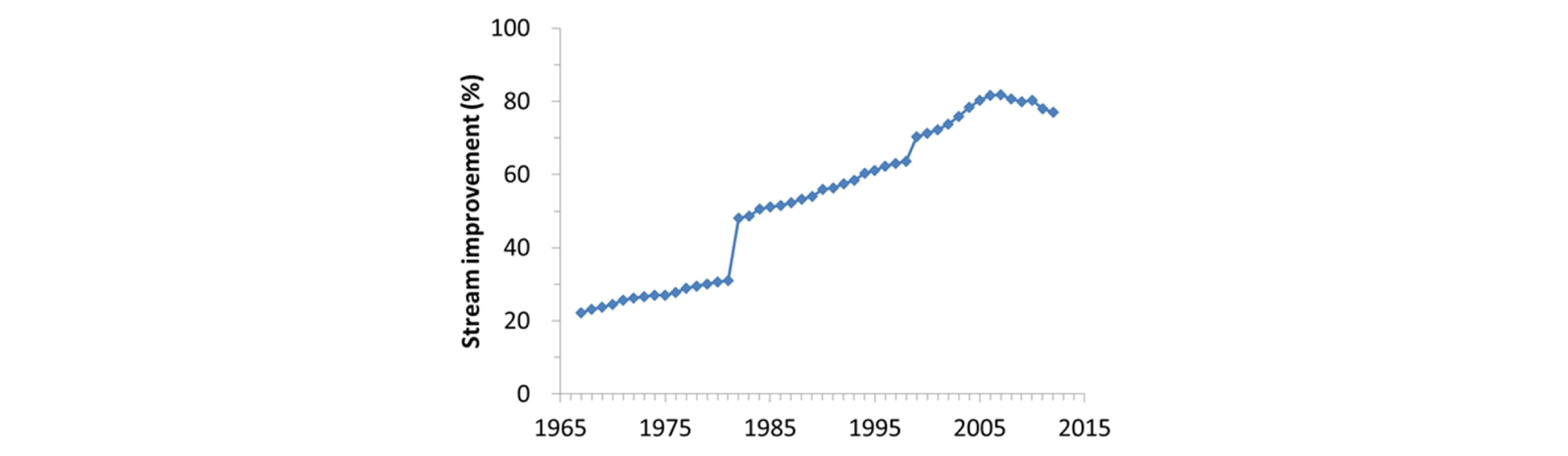
Fig. 1.
Changes of the stream improvement percentage calculated by ratio of the steam length with the levees to that requiring the levee construction in South Korea (MOLIT 2019).
2. 하천생물지형학의 이해
2.1 생물과 지형 상호작용의 시간적 변화
지형학자들은 중력, 물, 바람, 빙하와 같은 환경 인자가 지표면의 형태를 변화시키는데 주된 관심을 기울여왔다. 이러한 오랜 전통 속에서 미생물, 동물, 식물을 포함한 살아있는 유기체가 지형학적 현상에 기여하는 역할에 대한 연구는 상대적으로 낮은 비중을 차지하였다 (Corenblit et al. 2008, Stallins 2006). 그러나 오늘날 학계는 지구 상에 존재하는 거의 모든 지형형성작용이 생물의 영향을 받고 있는 것으로 판단하고 있다. 즉, 풍화, 침식, 운반, 퇴적, 심지어 지반의 융기와 침강에 이르기까지, 땅의 형태를 만들고 변화시키는 모든 작용에 있어 생물의 활동이 직 ‧ 간접적으로 개입되어 있다는 가설은 이미 현장에서의 실험과 컴퓨터 시뮬레이션을 통해 검증이 거듭되고 있다 (Dietrich and Perron 2006, Phillips 2009, 2016). 심지어 사막에서 모래가 바람에 날리고 퇴적되는 현상마저, 지의류와 조류, 균류, 박테리아 등으로 구성된 이른바 “생물 지각 (biological crust)”의 영향을 배제한 채 설명할 수 없다고 알려져 있다 (Naylor 2005, Viles 2008). 이러한 문제 의식 속에서 대두된 생물지형학은 하천의 역동성을 연구하는데 유용한 이론적 틀을 제공하는 핵심적인 학문 분야로 인식되고 있다 (Bendix and Hupp 2000, Osterkamp and Hupp 2010, Wohl 2013, Gurnell 2014).
하천생물지형학에서는 다양한 생물 중에서 특히 식물과 지형이 끊임 없는 상호 작용을 통해 하나의 생물지형 복합체 (biogeomorphic complex)를 구성한다고 알려져 있다 (Fig. 2). 예를 들면, 식생은 유수의 속도와 방향 등을 제어함으로써, 하천지형형성 작용에 간접적으로 개입할 수 있다. 또한 식생은 뿌리 조직으로 기질 (substrate)의 응집성 (cohesiveness)을 결정하고, 지표 상부 조직을 통해 물질의 침식, 운반 작용을 조절함으로써, 지표면 형태 변화의 규모와 속도에 직접적으로 영향을 준다. 이런 점에서 식생은 하천생물지형학에서 대표적인 생태계 기술자 (ecosystem engineer; Jones et al. 1994)라고 할 수 있다. 반대로 지형은 그 자체로서 서식처가 되어 다양한 토양 ‧ 수문 조건을 제공함으로써 식생의 생장과 분포에 직접적인 영향을 준다.
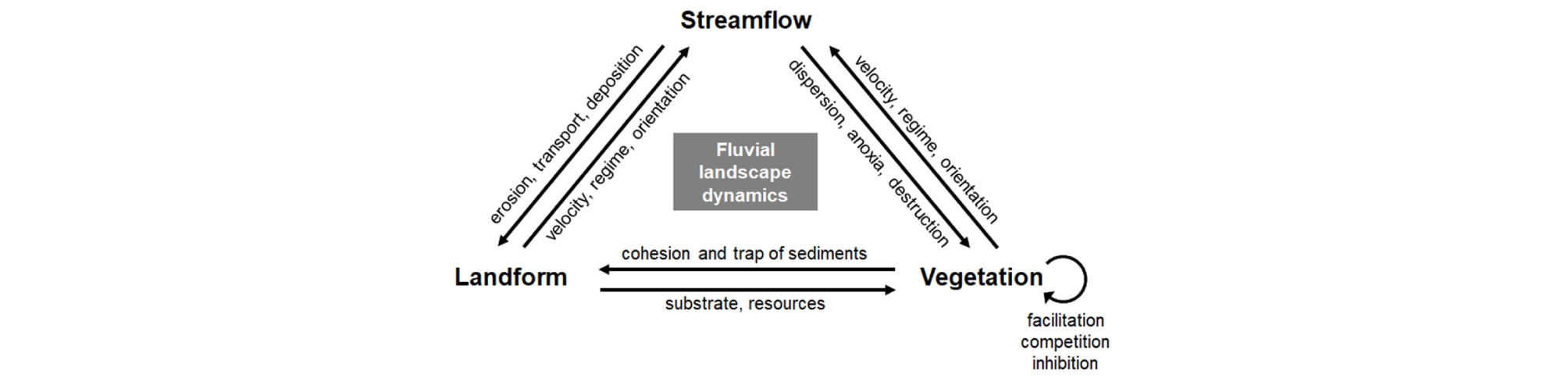
하천에서 생물과 수문지형 환경이 상호작용하는 생물지형 복합체는 다양한 시간과 공간 차원에서 변이를 가지게 된다. 먼저 시간 차원 측면에서 Corenblit et al. (2007, 2009)는 하천생물지형 천이 모델 (fluvial biogeomorphic succession model)을 제안한 바 있다 (Fig. 3). 이 모델은 크게 ‘지형적 (geomorphic)’, ‘선구 (pioneer)’, ‘생물지형적 (biogeomorphic)’, ‘생태적 (ecological)’ 단계로 나뉘어져 있다. 각 단계는 식생과 지형의 상호작용 측면에서 뚜렷한 특징을 지니고 있다. 첫 번째 단계는 강한 홍수가 기존의 식생피복과 기질을 대부분 제거한 후 시작되는 교란 직후의 상황을 의미한다 (Poole 2002). 그러므로 앞서 언급한 식생과 지형 사이의 긴밀한 상호작용은 거의 존재하지 않으며, 하상의 형태와 안정도는 기질 자체의 응집성과 수문적 조건에 의해 결정된다 (Brookes et al. 2000, Tabacchi et al. 2005). ‘선구’ 단계에서는 홍수에 의해 형성된 나지에 비로소 식물의 종자가 정착하고 발아하게 된다. 식생과 지형은 상호작용을 하기보다는, 식물종의 생존 ‧ 생장이 여전히 지형 ‧ 수문적 요소에 의해 좌우되는 일방적인 관계를 보인다. ‘생물지형적’ 단계에 이르게 되면, 식생은 비로소 유수의 속도와 방향, 기질의 침식, 물질의 운반 및 퇴적을 직접적으로 조절함으로써 하천지형형성작용에 주요한 인자가 된다 (Bendix and Hupp 2000, Brooks et al. 2003). 마지막으로 ‘생태적’ 단계에서는 기질이 하천의 수문지형학적 영향으로부터 어느 정도 단절되어 점차 안정되고, 식물종 사이의 경쟁에 의해 천이가 진행된다. 위 네 가지 단계는 홍수의 빈도와 강도에 따라 어느 시점에서든지 하천생물지형천이 모델의 최초 시작점으로 돌아갈 수 있다 (Baptist 2005, Geerling et al. 2006). 그 동안 이 모델은 내륙 하천 (Corenblit et al. 2010, Davies and Gibling 2013, Bätz et al. 2015), 염습지 갯골 (Kim 2012, Balke et al. 2014), 빙퇴석 (moraine; Eichel et al. 2013)을 포함하는 다양한 유형의 생태계를 이해하는데 적용되어왔다 (Corenblit et al. 2015).
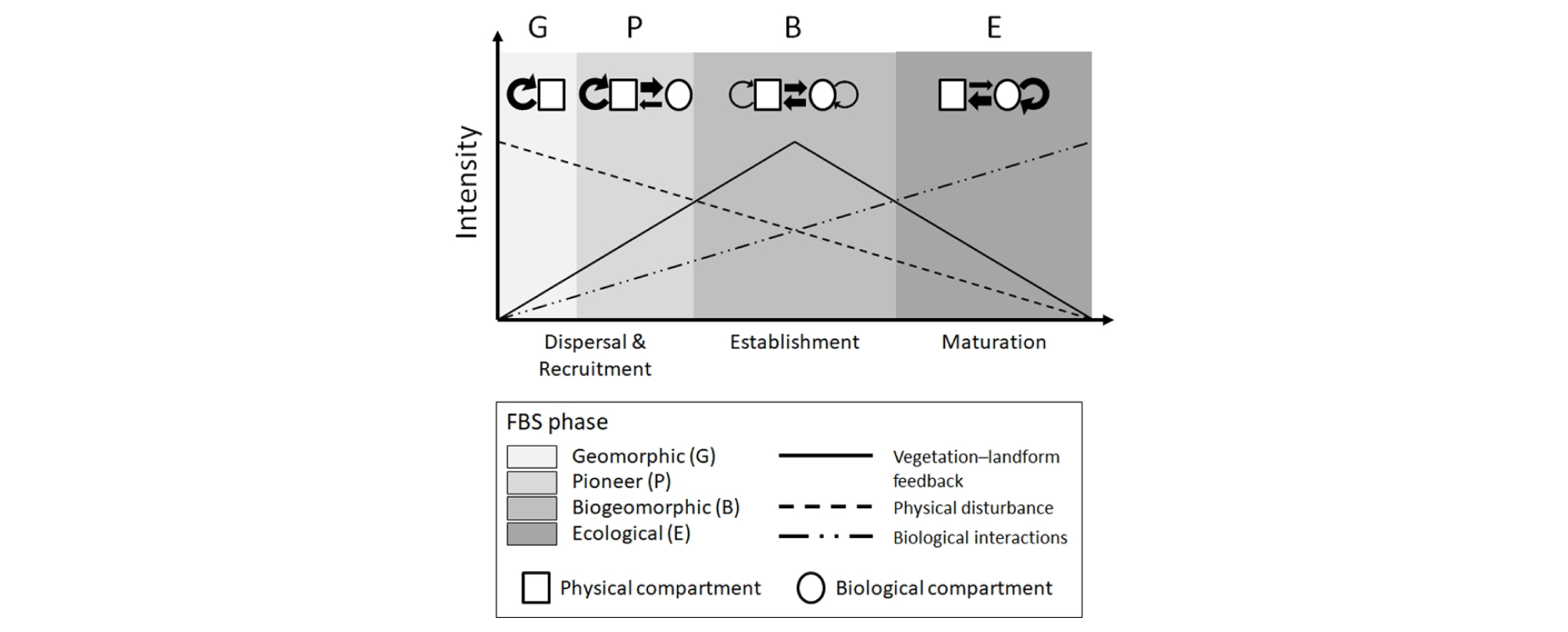
Fig. 3.
A simplified version of the fluvial biogeomorphic succession model first proposed by Corenblit et al. (2007, 2009). This is a modified version of Fig. 2 in Kim and Kupfer (2016).
2.2 생물과 지형 상호작용의 공간적 변이
하천에서 시간에 따라서 변화하는 생물지형 복합체는 공간 차원에서 이질성을 형성하게 된다. 특히 하도 식생은 흐름과 유사이송에 많은 영향을 주고 있으며, 지형변화에 중요한 역할을 한다. 하도 식생대에서는 흐름에 대한 저항이 증가하고 유속이 감소하며, 흐름 방향이 변화된다. 하도 식생에 의한 흐름 저항을 정량적으로 예측하기 위하여 많은 연구가 수행되어 왔는데, 이 저항은 식생의 크기, 강성, 물에 잠긴 정도, 밀도 등에 의하여 영향을 받는다 (Nepf and Vivoni 2000, Defina and Bixio 2005). 또한 반대 과정으로서 식생대와 주수로 사이에서 에너지가 교환되고 난류의 강도가 증가하며 주수로에서 흐름이 집중되어 하도가 변화하고 식생대에서 유사가 퇴적된다 (Tsujimoto 1999, Nepf and Vivoni 2000, Truong and Uijttewaal 2019). 하도 식생은 항력과 흐름 저항을 증가시키고 하상에 작용하는 전단응력을 감소시켜서 유사 이동이 감소되고 유사 퇴적이 증가되어 사주가 증가한다 (Thorne 1990, Crosato et al. 2012).
하천에서 사주는 하도 특성을 대표하는 중요한 형상으로서 흐름과 유사의 상호작용에 의하여 형성된다. 다양한 형상을 만드는 사주에서는 하도의 선형과 지형변화, 하도 단면의 확대, 하안 침식, 유사의 분급 현상이 나타난다. 하상의 침식과 퇴적 과정은 사주의 형성과 거동에 의해 영향을 받는다. 하도에 형성되는 사주는 하도의 횡방향으로 존재하는 사주가 1개인 교호사주 (alternate bar)와 2개 이상인 복렬사주 (braided bar)로 구별할 수 있다. 또한 사주의 이동 특성에 따라서 자유사주 (free bars)와 강제사주 (forced bars)로 구분할 수 있다. 자유사주는 하류로 이동하는 특성을 가지고 있으며, 강제사주 중에서 일반적으로 사행하천의 만곡부 안쪽에 발달한 고정사주 (point bars)는 거의 이동하지 않는다 (Blondeaux and Seminara 1985).
하도 식생은 하안의 안정성에 영향을 미친다. 식생의 밀도가 증가함에 따라 하안침식과 저수로의 이동이 감소하고 하도의 안정성이 증가한다 (Gran and Paola 2001, Jang and Shimizu 2007, Tal and Paola 2010). 이는 식생 뿌리가 견고하게 토양을 감싸고 있기 때문이다. 식생 뿌리가 감싸지 않은 흙은 압축력이 강하지만 인장력이 거의 없다. 식생 뿌리는 인장력과 탄성력을 증가시켜 응력을 분산시키는 역할을 한다. 또한 홍수터 식생은 강수를 차단하고 증발산에 의하여 토양수를 배제하여 토양건조를 촉진시키며 간극수압을 저하시켜서 하안의 안정성을 증가시킨다. 특히 하안에 있는 식생은 조도를 증가시켜서 유속과 흐름 에너지를 감소시키고 국부적으로 경계층 (boundary layer) 두께를 증가시키며 하안에서 전단응력을 감소시킨다 (Thorne 1990, Gran and Paola 2001). 결국 흐름이 하안에 직접 충돌하여 수충부를 형성하지 못하므로 2차류가 형성되어 하상에서 세굴이 발생할 수 있다 (Gran and Paola 2001). 또한 홍수터 식생에 의한 하도의 안정성 변화는 사주의 이동과 형상에 영향을 준다. 식생의 밀도가 높아져서 하안의 안정성이 증가하면 하폭 증가율이 감소하고 하폭 대 수심의 비가 감소한다. 또한 사주의 파장은 짧아지고 사주의 이동속도는 증가한다 (Jang and Shimizu 2007, Jang and Kim 2019, Vargas‐Luna et al. 2019). 이러한 사주의 변화는 결국 하천의 지형 변화에 큰 영향을 미친다.
하천 식생대에서는 유속이 느리고 저수로의 이동이 감소한다. 식생에 의하여 저수로가 고착되면 하상이 불규칙하게 변하면서 사주의 수가 증가한다. 식생의 밀도가 증가함에 따라서 유사 유출량이 크게 감소하고 저수로와 사주의 이동성을 나타내는 지표인 유사유출량의 변동성이 감소한다. 식생의 밀도가 증가함에 따라 망상하천에서는 저수로 수가 감소하거나 단일하천으로 변한다 (Fig. 4). 이때 저수로의 사행도와 하상의 기복지수는 증가한다 (Jang and Shimizu 2007, Jang 2013).

Fig. 4.
Temporal changes of the channel with vegetation: (a) run time (t) = 125 min, (b) t = 215 min and (c) t = 275 min. Arrows indicate the thalweg of the channel. The flow direction was from top to down (Jang and Shimizu 2007).
식생대에서 유속이 감소되고 유사가 퇴적되어 사주가 증가하고 유사 입경이 가늘어지면서 하천환경이 변한다. 이렇게 변화된 하천환경에서는 하도의 역동성이 떨어진다 (Jang and Shimizu 2007, Tal and Paola 2010). 하도 식생은 홍수 변화와 상류에서 공급되는 유사량의 변화에 영향을 받으며, 이러한 변화가 결국 하도 지형을 변화시키는데 중요한 역할을 한다 (Jang and Shimizu 2007). 하도 식생은 하상토의 유사 분급에 영향을 준다. 식생 밀도가 증가함에 따라 유사량은 감소하고 하상고는 불규칙하게 변하며 하상토 표층에서 유사 입경이 가늘어 진다. 식생대에서 유사가 포착되거나 식생대와 주 흐름 사이의 경계층에서 흐름이 발생하여 유사가 퇴적되고 식생대에서 흐름의 방향이 변화되어 표층에서 하상토 입도가 불규칙하게 분포한다. 또한 하상의 기복도가 증가함에 따라서 국부적으로 하상경사가 변하고 유사의 선택적 이동에 의하여 하상토가 분급되면서 평면적으로 입경이 불균일하게 분포하는 분급 특성이 나타난다 (Jang 2016). 결국 하도 형상과 식생에 의하여 유로 (flow channel)가 정해지고 유사 이송은 유로 형상과 식생에 의하여 영향을 받는다. 또한 하상토 분포와 유사 이송은 지형 변화에 영향을 미치면서 상호작용 체계가 형성된다 (Gurnell 2014). 유량 변화와 상류에서 공급되는 토사는 이 체계에서 상호작용을 형성하는데 중요한 경계조건이다. 하도 변화는 그 공간에서 흐름, 유사 분포, 식생 등이 상호작용하여 나타나는 과정이다. 그러므로 하천을 합리적으로 관리하기 위해서 하도 변화와 이들의 상호작용 과정을 파악하는 것이 중요하고 식생의 변화와의 상호관련성을 파악하는 것도 필요하다.
2.3 수문지형의 이질성과 생물다양성의 관계
앞에서 살펴본 하천에서 수문지형에 의한 서식처의 이질성은 그곳에 서식하는 생물다양성에 큰 영향을 미친다. 사행하천에서 만곡부의 내측에는 고정사주가 발달하며 외측에는 수심이 깊은 웅덩이가 발달하여 여울과 소 구조가 형성된다. 물의 흐름이 빠르고 수심이 얕은 여울 구간은 조립질의 유사가 퇴적되어 하상의 공극률이 높고 용존산소량이 상대적으로 많아서 저서성 대형무척추동물이나 어류의 산란 장소로 이용될 수 있다. 여울이 형성되는 지점에는 고정사주가 발달하는 경우가 많은데, 홍수기에 사주 위로 물 흐름이 집중되면서 지형 조건이나 식생 영향으로 세굴이 발생한다 (Brierley and Fryirs 2005). 홍수 이후 수위가 낮아졌을 때 사주의 상류 쪽 머리나 하류 쪽 꼬리 부분에 물이 정체되는 정수역이나 웅덩이가 형성된다. 일부 어류에서 산란은 여울에서, 휴식하거나 치어기에는 정수역에서, 성장 후에는 소에서 서식하기 때문에, 서식처 다양성은 생물다양성과 밀접한 관계를 가진다. 또한, 하루살이는 산란을 위해 본류의 여울이나 자갈사주를 이용하고, 성충이 되었을 때는 사주 위나 주변 식생에서 생활한다 (Yuma and Hori 1990, Holomuzki and Messier 1993). 이처럼 서식처의 이질성과 다양성이 생태적 지위를 확대하여 종다양성을 증가시킨다는 서식처 이질성 가설 (Simpson 1949)은 다양한 연구에서 증명되고 있다. 예를 들면, 일본의 Kizu 강에서 여울, 소, 사주 머리 정수역, 사주 꼬리 정수역, 모래사주 위 웅덩이 (active pond), 식생사주 위 웅덩이 (terrace pond)에서 저서대형무척추동물의 종다양성은 서식처 다양성과 밀접한 관계를 보였다 (Choi et al. 2012) (Fig. 5).
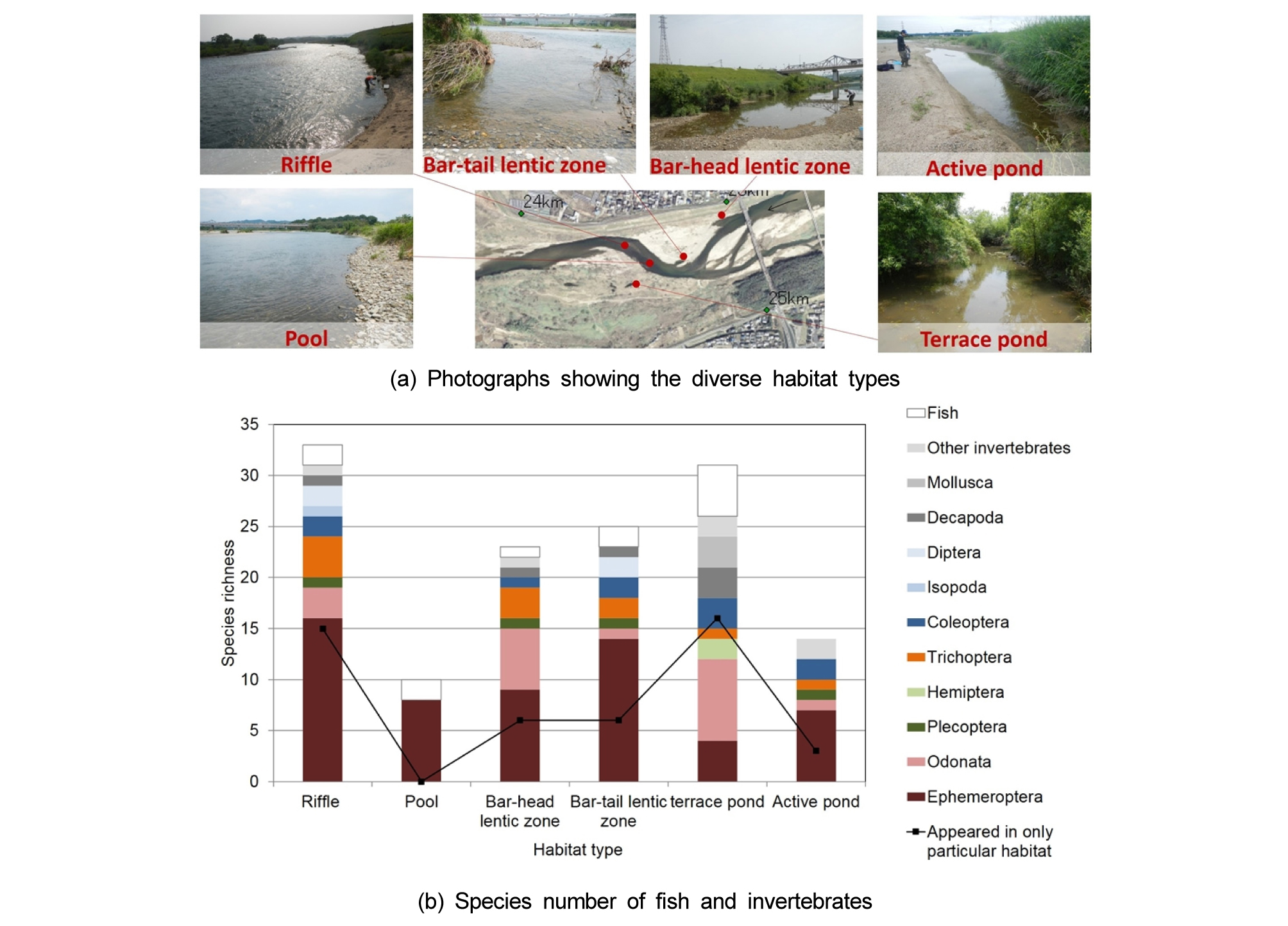
Fig. 5.
Comparisons of species richness of macroinvertebrates with the different habitat types in the Kizu River, Japan (Choi et al. 2012).
2.4 교란과 생물지형 복합체의 관계
유역에서 물 흐름과 유사 이동 사이의 에너지 균형과 상호작용에 의하여 하천 지형이 형성되며 (Leopold and Wolman 1957), 상류에서 하류까지 침식, 이동, 퇴적 작용을 반복하면서 여울과 소, 사주와 같은 하도의 서식처 구조가 형성되고 유지된다 (Lane and Richards 1997). 유역 차원에서 하천에 가해지는 가장 크고 광범위한 인위적 교란은 하천을 횡적으로 가로질러 건설된 댐, 대형보, 하구둑과 같은 대형 수리구조물 (이후 댐으로 통칭함)이라고 할 수 있다 (Rosenberg et al. 2000). 이러한 수리구조물은 물 흐름과 유사 이동의 각각에 또는 동시에 영향을 주기 때문에, 댐 건설에 따른 인위적인 유황 변화와 유사 이동의 단절은 하천 지형에 구조적인 변화를 가져오게 된다 (Petts and Gurnell 2005). 그리고 댐 하류 조절하천에서 이러한 생물서식처의 물리적 변화는 연쇄적으로 이를 기반으로 살아가는 어류, 저서생물, 조류 등의 생물종다양성을 감소시키고 생태계 물질순환에 영향을 주고 있다 (Power et al. 1996, Ock and Takemon 2014).
먼저 조절하천에서 유황변화에 따른 교란을 살펴 보면, 댐 조성에 의하여 저수지에 물을 저장하면서 시간적으로 댐 건설 전 ‧ 후, 그리고 공간적으로 상 ‧ 하류 간에 수문학적 변화가 발생한다 (Richter et al. 1998, Magilligan and Nislow 2005). 다목적 댐을 예로 들면, 상수원수, 농업용수 등으로 물을 취수하는 만큼 하류로 방류되는 유량의 절대량이 감소되어 댐 하류에서 평수위가 낮아진다. 홍수기에는 홍수 조절로 인하여 첨두유량이 감소하고 홍수위의 빈도가 줄어든다. 그리고, 갈수기에는 발전을 위하여 일정 유량의 방류가 지속되면서 댐 조성 이전보다 갈수위가 높아지게 된다. 따라서 댐 하류 조절하천에서는 홍수량과 홍수 빈도가 감소되고 갈수위가 상승하여 하천유량이 연중 단순화 및 균일화되어 상하류 간에 유황이 급격히 변한다.
다음으로 조절하천에서 유사이동 단절에 따른 교란을 살펴보면, 일반적으로 댐 건설에 의해 저수지가 만들어지면 자갈과 모래와 같은 큰 입자의 소류사는 하류로의 이동이 단절되며, 특히 대규모 저수지에서는 부유사를 포함하여 99% 이상의 유사이동이 차단된다는 보고가 있다 (Williams and Wolman 1984). 반면에, 댐 하류로는 세류사라고 부르는 미사와 점토와 같은 미세립자만이 이동하게 된다. 이 세류사는 하상에서 모래와 자갈 사이의 틈을 메우면서 물과 산소의 이동을 방해하여 하상이 혐기화되고 저서생물의 서식이 저해되는 이차 피해를 가져온다.
댐에 의한 유사이동 차단과 유사공급의 불균형은, 댐 저수지에서는 퇴사로 인하여 댐의 유효저수용량을 감소시키고 댐 하류역에서는 유사공급량이 감소되어 유사 부족의 문제를 발생시킨다. 이러한 유사가 결핍된 유량 (sediment starvation flow)은 제방과 강바닥을 침식시켜 유사 농도를 맞추려고 하기 때문에 하도의 세굴력을 증가시켜 하상 저하를 유발하고 하상 재료의 조립화를 가져온다 (Graf 2006). 그리고, 이러한 영향은 저수지에서 강, 바다까지 그 영향이 지속적이고, 광범위하게 발생하기 때문에, Kondolf (1997)는 ‘hungry water’ 라는 개념으로서 댐 하류 조절하천에서 발생하는 하천 구조물의 교각 세굴, 여울과 소와 같은 생물서식처 소실, 해안선 후퇴와 같은 지형 변화를 과학적으로 설명하였다.
3. 우리나라 하천에서 수문지형과 생물 교란
3.1 식생 번무화
최근 우리나라의 많은 하천에서 식생이 과도하게 발생하는 경우가 있다. 과거에는 주로 댐 하류 조절하천에서 댐의 유황 조절로 인하여 식생이 번성하는 것이 일반적이었다 (Woo 2008). 최근에는 댐 유무, 하천 크기, 지역 등에 관계없이 전국적으로 하천에 식생이 과도하게 발생하고 있다. 섬강과 내성천에서는 조사구간 하천 면적의 약 50% 이상을 식생이 덮고 있으며, 청미천에서는 하천 면적의 78%를 식생이 차지하고 있었다 (Kim and Kim 2019). 식생 도입 속도가 매우 빨라서 내성천에서는 2011년부터 2018년까지 식생 면적이 17배 증가하였다. 전국적으로는 살펴보면 우리나라 국가하천의 약 34%가 식생에 의해 잠식되어 있었다 (KICT 2015).
이와 같이 최근 비조절 하천에서 식생이 급증하는 원인을 여러 가지로 추정할 수 있다. 먼저 강우 발생 유형의 변화로 식생 발아기에 과거와 같이 강우가 자주 발생하지 않고 있다. 두 번째는 하도에서 골재 채취, 하천정비 등의 인위적 교란이 식생 이입을 촉진하고, 세 번째는 하천에 영양물질 유입량이 증가하여 식물 생장이 촉진되고 있다고 추측된다 (Woo and Park 2016). 이밖에 산지의 수림화로 인하여 하천으로의 유사 유입이 줄어든 것도 또 다른 원인으로 추정되고 있다.
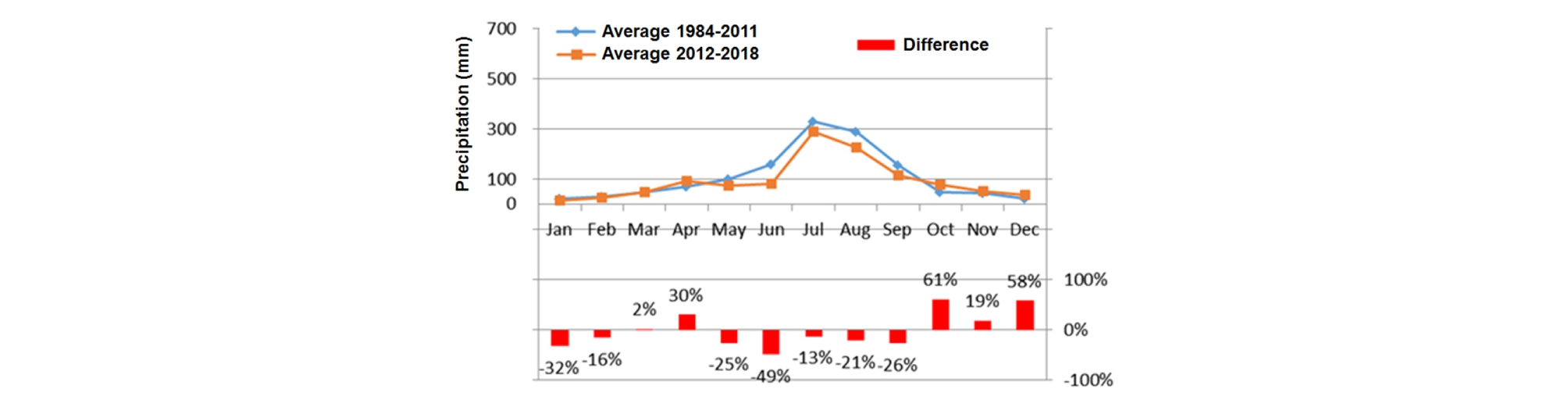
남한의 전역에 걸쳐 하천의 규모와 위치에 관계없이 하천에 식생이 광범위하게 발생하고 있다는 점은 기후 요인 중에 하나인 강우 발생 패턴 변화가 이러한 변화의 주요한 요인이 될 수 있다는 것을 암시한다. 최근 연구 결과에 의하면 1984 - 2011년 (28년)과 2012 - 2018년 (7년)에 월강우량 유형이 크게 변화되었다 (Fig. 6). 즉 4월 강우량은 30% 증가하였고 5 - 9월까지의 강우량은 최대 48% 감소하였다. 이와 같이 5월 이후의 강우량 감소는 식생 발아기와 유식물 성장기에 큰 홍수 교란을 감소시켜 식생의 초기발생을 촉진할 수 있다. 특히 여름철 강우 감소는 발아된 유식물이 활착할 수 있는 조건을 제공할 수 있다. 우리나라에서 이와 같은 강우 패턴 변화에 따른 식생의 과도한 발생은 일본에서도 비슷하게 나타나고 있어서 하천의 식생 번무화가 우리나라에 한정된 국지적인 현상이 아니라고 생각된다.
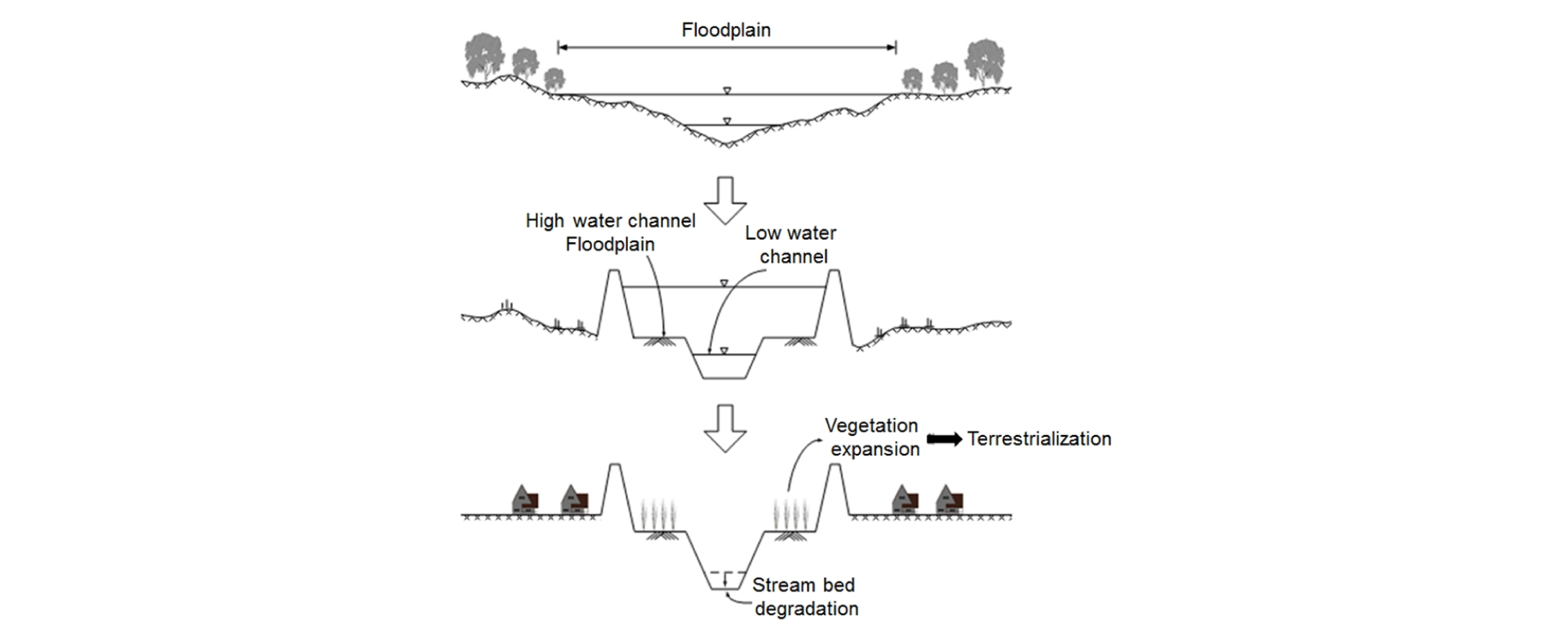
우리나라에서 하천 정비 정책도 식생 발생에 큰 영향을 미친 것으로 추정된다. 유황계수가 큰 우리나라에서는 하천을 정비하면서 흔히 복단면 하도로 조성하였는데, 이러한 하천 구조가 하천에 식생 발생이 가중되는 하나의 원인이 될 수 있다고 생각된다 (Fig. 7). 복단면 하도에서는 저수로에 물이 집중되고 고수부지에 물이 흐를 기회가 줄어들어서 식생이 과도하게 발생할 수 있는 환경이 형성될 수 있다. 일단 식생이 정착한 후에는 고수부지 식생으로 인해 저수로에서 유속이 빨라지고 세굴이 발생하여 저수로 저하현상이 발생한다. 이 결과로서 또 다시 고수부지에 물 흐름의 기회가 상실되는 결과를 낳아서 이러한 과정이 심화 반복하게 된다. 결국 하천의 복단면 구조로 인하여 고수부지에 식생이 정착하게 되고 저수로와 고수부지의 단차가 증가하면서 고수부지에 식생이 활착하는 조건이 형성될 수 있다.
식생의 과도한 발생은 홍수소통의 단면을 감소시키고 하도를 육역화하기 때문에 하천 관리와 생태계 보전에 부정적인 영향을 미칠 수 있다. 따라서 식생 과도한 번무화에 대한 원인 분석과 대책 마련이 필요한 시기라고 생각된다. 기후변화로 인한 현상이라면 적응관리를 포함한 다양한 대책을 수립하여야 하고, 복단면 하도 조성으로 발생하는 식생을 관리하기 위해서는 단단면 조성으로 하천관리 정책을 개선하는 것을 적극적으로 고려할 필요가 있다.
3.2 외래식물 침입
수생태계와 육상생태계의 경계인 하천 주변 지역은 물의 흐름으로 인한 범람이나 가뭄, 퇴적 등이 지속적으로 일어나면서, 주변 식물 번식체의 유입이 활발하고 교란이나 환경 조건의 변화가 높은 빈도로 발생하는 매우 역동적인 생태계이다. 이러한 특성으로 인해 비교적 좁은 공간 내에서 높은 종 다양성을 보이며, 홍수의 조절, 영양분 및 퇴적물 보유, 수질 개선 등 다양한 생태계 서비스를 제공한다 (Richardson et al. 2007). 반면 하천변의 이러한 물리적 특성은, 수변 지역의 인위적인 교란 (인공호의 조성, 유량의 조절, 제방이나 다리 건설, 식생 정비 등)과 함께 외래식물의 유입과 정착을 활발하게 만드는 요인이기도 하다 (Pattison et al. 2019). 유입된 외래식물은 기존 생태계를 변화시킬 수 있는데, 예를 들면 식물 생태계의 종 다양성 감소, 먹이 사슬 변화, 탄소나 질소와 같은 양분 순환의 변동 등을 야기하고, 그 결과로 수변 생태계의 생태계 서비스를 감소시킬 수 있다 (Zedler and Kercher 2004).
외래식물종의 침입에 따른 식물 생태계의 변화는 수문지형 또한 변화시키는 것으로 알려져 있다 (Fei et al. 2014). 식물은 광합성 과정에서 토양 수분을 공기 중으로 증발산시키는데, 생태계 구성 식물종의 변화는 토양 수분의 발산 정도를 변화시키면서 수문학적 변화를 야기할 수 있다. 예를 들면 미국 서부지역에 침입한 Tamarix 종은 활발한 증발산 활동으로 인해 강폭을 감소시키고 토양의 염도를 증가시키는 것으로 알려져 있다 (Zavaleta 2000). 또한 외래식물종의 침입으로 인한 생태계 내부 식물종의 뿌리나 줄기의 변화는 퇴적과 침식의 형태를 변화시킬 수 있고 이로 인해 지형이 바뀔 수도 있다 (Crooks 2002).
외래식물종의 확산이 야기하는 다양한 문제의 해결을 위해 외래식물종 침입 및 확산의 생태학적 기작에 관한 연구가 지속적으로 수행되어 왔다. 연구 결과들을 종합해 보면, 외래식물종의 침입 가능성은 교란의 정도나 양분과 같은 비생물학적 환경 조건과 함께, 생태계를 구성하는 식물종의 특성에 영향을 받는다 (Catford et al. 2009, Byun et al. 2018). 예를 들면 외래식물종 침입 이전에 자생하는 식물종 중 일부가 외래식물종과 유사한 생태적 지위를 갖거나, 생태계 우점종이 외래식물종 대비 경쟁 우위를 갖는 경우 외래식물종의 침입 가능성이 감소할 수 있다. 혹은 생태계의 종 다양성이 높아질수록, 특히 생태계에 존재하는 기능군 (functional group)이 다양할수록 외래식물종의 침입이 어려워진다. 즉 식물 생태계의 특성에 따라 외래식물종에 대한 저항성이 상이하게 나타난다.
최근에는 이러한 외래식물종 침입의 생태학적 기작을 이용하여, 외래식물종의 침입을 막을 수 있는 종 구성을 지닌 생태계를 복원하고자 하는 생태공학적 접근 방법에 대한 연구들이 진행 중이다 (Schuster et al. 2018). 외래식물종의 관리는 전통적으로 물리적인 제거, 제초제 등을 이용한 화학적 제거, 천적을 이용한 생물학적 제거의 방법이 이용되어 왔다. 그러나 성공적으로 정착한 대부분의 외래식물종들은 많은 양의 씨앗을 생성하여 토양에 다량의 매토종자를 남긴다. 물리 혹은 화학적 제거 방법이 적용된 구역이 후속 조치 없이 방치될 경우, 제거 자체가 교란 인자로 작용하여 외래식물종이 매토종자로부터 다시 발아, 정착할 가능성이 높다. 만약 외래식물종의 제거 이후 외래식물종의 정착 가능성이 낮은 생태계를 복원할 수 있다면 외래식물종의 재확산을 보다 효율적으로 방지할 수 있을 것이다. 이러한 방법을 이용하기 위해서는 외래식물종의 침입성과 생태계 구성 식물종들 사이의 인과관계에 대한 연구가 필수적인데, 수목이나 초지 생태계에 비해 수변 생태계를 대상으로 하는 연구는 아직 미흡한 실정이다 (Pattison et al. 2019).
외국의 사례와 유사하게, 우리나라의 하천 주변에도 많은 외래식물이 침입한 것으로 보고되고 있다. 대표적 외래식물종으로는 미국좀부처꽃 (Ammannia coccinea)과 털물참새피 (Paspalum distichum var. indutum), 가시박 (Sicyos angulatus), 돼지풀 (Ambrosia artemisiifolia)이 있다. 이와 함께 환삼덩굴 (Humulus japonicus)은 자생종이지만 지속적인 서식지 확장과 침입 생태계에 대한 부정적 영향으로 인해 최근 생태계교란생물로 지정되었다 (Information of Korean Alien Species 2019). 이들 식물종들의 분포나 생태계에 주는 영향에 대한 연구들이 국내에서 지속적으로 수행 중이다 (Oh et al. 2008, Jin et al. 2013, Hwang et al. 2014, Cho and Lee 2015, Lee et al. 2015, Lee et al. 2016). 그러나 외래식물종으로 인한 식물 생태계의 변화가 수문지형에 줄 수 있는 영향은 간과되고 있는 실정이다. 예를 들면 미국좀부처꽃과 털물참새피의 경우 줄기가 물에 잠긴 상태에서 왕성한 생육을 보이고, 이들의 뿌리와 줄기는 물의 흐름과 퇴적에 영향을 줄 수 있을 것이다. 가시박이나 환삼덩굴은 타 하천 식물종들에 비해 빠른 생육을 보이는데, 이는 토양 수분의 증발산량을 증가시켜 수문학적 변화를 야기할 수 있다. 외국의 사례를 참고로, 국내의 사례에 맞는 수문지형학적 연구가 필요하다.
우리나라의 하천에서는 1960년대 하천법이 제정된 후 치수를 목적으로 한 광범위한 인위적 교란이 지속적으로 진행되어 왔다. 이러한 교란으로 인해 외래식물종이 광범위하게 하천 생태계에 침입했을 것으로 추정된다. 일례로 Kang et al. (2009)은 2005년부터 1년간 진행된 우포늪 제방 공사 이후, 이전에 없었던 환삼덩굴과 돼지풀의 출현을 보고하였다. 그러나 앞에서 언급한 생태계 구성 식물종에 따른 외래식물종에 대한 저항성을 고려하면, 침입 이후 외래식물종의 정착과 확산은 교란 이후 복원 방법에 영향을 받을 것이다. 사업직후 초기식생 정착 기간에 환삼덩굴이 침입하여 우점하였지만, 시간이 경과됨에 따라 갈대 군집으로 대체되는 양상이 보고된 양재천 복원사업의 결과 (Cho et al. 2008, Lee et al. 2011)는 시사하는 바가 크다고 하겠다. 많은 자원의 투입이 요구되는 물리적, 화학적 제거 방법에 비해 생태계의 저항성을 이용하는 방법은 효율성이 매우 높을 것으로 기대된다. 독특한 국내 하천 생태계의 특성과 저항성을 고려한 외래식물종 제어 방안 마련을 위한 연구가 필요한 시점이다.
4. 결론 및 전망
하천생태계는 식생을 포함한 생물과 지형, 토양, 수문 등의 다양한 환경요인이 긴밀한 상호관계를 유지하며 시공간에 따라서 역동적으로 변하는 다중복합체이다. 이와 같은 개념은 하천을 연구하고 관리, 복원 기술을 개발하는데 두 가지 의미를 갖게 한다. 첫째, 위의 환경요인 각각에 대한 단편적인 연구와 지식으로는 하천 시스템에 대한 전체론적 이해와 관리를 도모하기 어렵다. 요인 사이의 되먹임이 발생하고 소멸하는 과정에 대하여 구체적으로 이해하기 위한 꾸준한 노력이 필요하다. 둘째, 홍수와 같은 대규모 교란 직후 또는 이를 대비하기 위한 선제적 전략을 수립할 때에 어느 한두 요인의 유지와 관리에 국한된 접근은 큰 성공을 거두기 어려울 것이다. 예를 들어, 식생을 관리, 복원하려면 적절한 토양과 지형 조건이 무엇이냐에 대한 충분한 고민을 하여야 한다. 반대로 하도 내에 특정 지형 경관을 유지하기 위해서는 어떤 종의 식물을 어떤 방식 (밀도, 배치 등)으로 식재할 것이며, 이들의 존속을 위해 어떤 토양 성질을 조성할 것인지 등에 대한 고민이 수반되어야 한다.
생물에 의하여 역동적인 영향을 받는 하천의 다중복합체에서 특히 식생에 의한 지형변화 연구가 활발하게 진행되어 왔다. 그러나 식생에 의한 하도 지형변화를 정량적으로 예측하여 하천관리를 하기에는 아직 한계가 있다. 따라서 식생에 의한 흐름의 저항, 식생대에서 흐름 특성 및 운동량 교환을 정량적으로 파악하고, 하도 지형변화를 예측할 수 있는 정교한 수치모형을 개발하는 것이 필요하다. 또한 하도의 지형 변화와 식생의 상호관련성을 정량적으로 파악하여, 홍수에 안전하고 건전하고 경관과 어울리는 하천으로 유도하기 위하여 하도 식생 관리 지침을 제시하는 것도 필요하다.
현재 하천에는 인간에 의한 다양한 교란이 가해지고 있다. 예를 들면, 조절하천에서 발생하는 서식처 지형 변화와 생태계 변화는 댐을 보유한 국가에서 공통적으로 발생하는 국제적인 현상이다. 더불어, 신규 댐 건설의 한계와 기존 댐의 노후화로 인하여 선진 각국에서는 조절하천의 자연성 회복을 21세기 하천관리 목표로 설정하고 있으며, 이와 관련한 연구와 기술은 우리의 예상보다 빠르게 발전하고 있다 (Ock et al. 2019). 최근 우리나라의 하천에서 최근 가장 눈에 띄는 생태계 변화에 식생이 과도하게 발생하는 것과 외래식물이 침입하는 것이 속한다.
우리나라 하천의 규모, 위치, 유량조절 여부 등을 떠나 전국적으로 거의 모든 하천에서 식생이 과도하게 발생하고 있다. 식생 이입이 지속되면서 초본류 뿐만 아니라 목본류가 하천의 넓은 면적을 차지하고 있다. 이러한 식생의 과다 발생은 유역에서의 영양염류 과다 공급, 산지 수림화로 인한 유사 발생 감소, 댐으로 인한 유황 변화 등의 원인도 있지만 단기적 기후변화로 인한 강우발생 유형의 변화가 주요한 원인인 것으로 판단된다. 식생의 과도한 발생은 하천 통수단면적 감소, 하천 육역화, 하천 생태계 변화 등의 문제를 유발하여 일정 수준 이상으로 식생이 발생하면 하천관리 측면에서 부정적인 영향을 발생시킬 수 밖에 없다. 하천의 지속적인 기능 유지를 위해서는 하천의 물리적 공간, 하천 지형, 유황 등에 대한 분석을 통해 식생의 영향을 최소화할 수 있는 방안을 개발하는 것이 시급한 것으로 판단된다.
하천 주변은 자연적, 인위적 교란이 높은 빈도로 광범위하게 발생하는 공간으로 외래식물의 침입 및 정착이 매우 활발한 생태계이다. 이러한 특성으로 인해, 외래식물종의 정착 및 확산은 하천 식생 생태계의 생태계 서비스 효과나 수문지형에 대한 영향을 연구할 때 반드시 고려되어야 할 요소로 판단된다. 외국에서는 외래식물의 생태학적 특성에 기반하여 이들의 침입을 방지하거나 수문지형에 대한 영향을 최소화하기 위한 다양한 생태공학적 접근이 시도되고 있으나, 국내의 연구는 아직 초기단계라 할 수 있다. 외국 하천과는 상이한 국내 하천의 독특한 환경 특성을 고려할 때, 국내 하천의 환경에서 확산되고 있는 외래종을 대상으로 한 연구가 필요한 시점이다.
하천에 발생하는 교란에 대응하여 국내의 하천복원사업에서 식물을 식재하고 습지, 여울 등과 같은 서식처를 조성하여 생물다양성을 증진하고자 노력하여 왔다. 그러나 시간이 경과함에 따라 상류역의 수문지형적 조건이 변화하면서, 식생이 과도하게 활착하여 통수능이 저하되고, 토사 퇴적으로 인하여 습지가 매몰되거나 여울의 입경이 세립화 되기도 하는 등 서식처 환경이 변화하여 꾸준한 적응관리가 필요하다 (Choi et al. 2015). 결국 생물의 종다양성을 높이거나 특정 생물의 서식을 유도하기 위해서는 하천 지형과 서식처의 자연적인 변화와 이동을 고려할 수 있도록 중규모 하천지형과 유역 수준의 수리 지형적 특징까지 파악하는 체계적이고 종합적인 접근이 필요하다. 이를 위하여 하천의 조사 결과를 바탕으로 인과관계를 분석하고 관리방안을 도출하려는 노력이 필요하다고 판단된다. 특히 우리나라에서는 2018년 6월에 물관리일원화법이 통과되면서 하천 자연성 회복을 위한 수량과 유사, 수질, 생태계의 통합관리 기반이 마련되었다. 따라서 앞으로 통합관리체계를 기반으로 하는 수생태계 자연성 회복과 관리를 위한 하천생물지형학 연구가 보다 활성화되기를 기대한다.
이제 하천생물지형학자들은 다양한 요인들 사이에 발생하는 상호작용을 정량화하는 문제에 대해 고민할 필요가 있다. 우리는 이미 지형이 식생의 분포에 영향을 준다는 일방향적인 인식에서 벗어나, 식생이 능동적으로 지형 형성 프로세스를 제어한다는 양방향적인 사고를 하고 있다. 마찬가지로 우리는 토양 조건이 식생의 생장에 영향을 줌과 동시에, 식생 역시 뿌리 조직을 통해 토양 기질을 고정하고 지화학적인 영향을 주고 있음을 알고 있다. 그렇다면 삼자 사이의 관계를 살펴봄에 있어, 식생은 종속변수가 되고 지형 ‧ 토양 인자는 독립변수가 되는 전통적인 이분법적 회귀분석의 틀에서 벗어나야 할 필요가 있다. 현재 하천생물지형학은 방법론적 발전이 이론적 발전을 충분히 따라가지 못하는 상황이라고 진단할 수 있다. 한 가지 방법으로 식생과 토양, 지형 (또는 수문) 삼자 사이의 관계를 벤 다이어그램으로 시각화하는 것을 상상해볼 수 있을 것이다 (Kim et al. 2015, Kim and Kupfer 2016).
몬순 기후의 우리나라 하천은 연중 유황 변화가 큰 높은 하상계수를 갖고 짧은 거리에서 산지로부터 바다까지 하천이 급경사로 흐르는 매우 독특한 수문지형 환경을 갖고 있다. 또한 인구밀도가 매우 높아서 하천에 인접한 곳까지 토지를 이용함으로써 홍수재해를 방지하기 위하여 하천 지형과 수문 조건을 변경하고 생물을 교란하여 왔다. 그러나 하천의 생물다양성을 확보하고 생태계 서비스를 보장하고자 하는 요구가 경제 발전과 함께 높아지고 있다. 하천 생태계를 친자연적으로 관리하고 복원하기 위해서는 생물지형학적 관점에서 생물과 환경 사이의 상호작용을 양방향으로 이해하는 노력이 필요할 것으로 판단된다.
